Interactome Networks!
Using programs such as Osprey and SPRING, I found the interactome networks Sox9 belongs to in mice and humans (SPRING 8.0), and just in humans for Osprey. Many of the interactions have overlap in both mice and humans, with some being mentioned frequently in the literature of Sox9's role in endochondral bone development. Many of the SPRING interactions are indirect. This is due to Sox9 being a transcription factor, and would not need to have physical interactions with other proteins to genetically interact with them. However, Sox9 was indeed shown to physically bind to Runx2, Col2a1, and Nr5a1 in both mouse and human; Creb1, Med12 are found in the human network only, and Ctnnb1 is only in the mouse network. This discrepancy can either be true species specific interacters of Sox9, or more likely, interactions that exist in both, just not have been demonstrated yet in the respective literature of each organism.
In the Osprey database, there were interesting physical interactions found between Sox9 and MAF, MED12, NR5A1, and HSP1A1. In particular, the MAF and NR5A1 interactions were the most interesting, not due to themselves but to their other interactions, which would be indirect/downstream/upstream interactions of Sox9. MAF is able to interact with HOXD12, CREBBP, and EP300. All of these are known important transcription factors involved in embryogenesis (Davis et al 1996; Zakany et al 1996; Kioussi et al 2002; Huelsken et al 2000; Shikama et al 2003; Elliot et al 2007). With NR5A1, it is shown to interact with Pitx1, which is a transcription factor that is essential for hindlimb development (Lanctot et al 1999). However, since NR5A1 (aka SF-1) is found primarily in sexual development and interacts with Sox9 in this process (De Santa Barbara et al 1998), and the fact that Pitx1 is also expressed in the pituitary gland where NR5A1 interacts with it (Tremblay et al 1999) and is found expressed in the hindlimb considerably before Sox9 expression, this interaction tree may be representative of Sox9's role in sexual development, not chondrogenesis. However, one should note that Ctnnb1 (Beta-Catenin) is indeed important for skeletal development (Day et al 2005; Hill et al 2005) so that interaction should not go unnoticed (especially since it is found in the other interactome data sets).
None the less, these interactors are all compelling representatives that should be looked into further, if they have not been extensively studied already.
In the Osprey database, there were interesting physical interactions found between Sox9 and MAF, MED12, NR5A1, and HSP1A1. In particular, the MAF and NR5A1 interactions were the most interesting, not due to themselves but to their other interactions, which would be indirect/downstream/upstream interactions of Sox9. MAF is able to interact with HOXD12, CREBBP, and EP300. All of these are known important transcription factors involved in embryogenesis (Davis et al 1996; Zakany et al 1996; Kioussi et al 2002; Huelsken et al 2000; Shikama et al 2003; Elliot et al 2007). With NR5A1, it is shown to interact with Pitx1, which is a transcription factor that is essential for hindlimb development (Lanctot et al 1999). However, since NR5A1 (aka SF-1) is found primarily in sexual development and interacts with Sox9 in this process (De Santa Barbara et al 1998), and the fact that Pitx1 is also expressed in the pituitary gland where NR5A1 interacts with it (Tremblay et al 1999) and is found expressed in the hindlimb considerably before Sox9 expression, this interaction tree may be representative of Sox9's role in sexual development, not chondrogenesis. However, one should note that Ctnnb1 (Beta-Catenin) is indeed important for skeletal development (Day et al 2005; Hill et al 2005) so that interaction should not go unnoticed (especially since it is found in the other interactome data sets).
None the less, these interactors are all compelling representatives that should be looked into further, if they have not been extensively studied already.
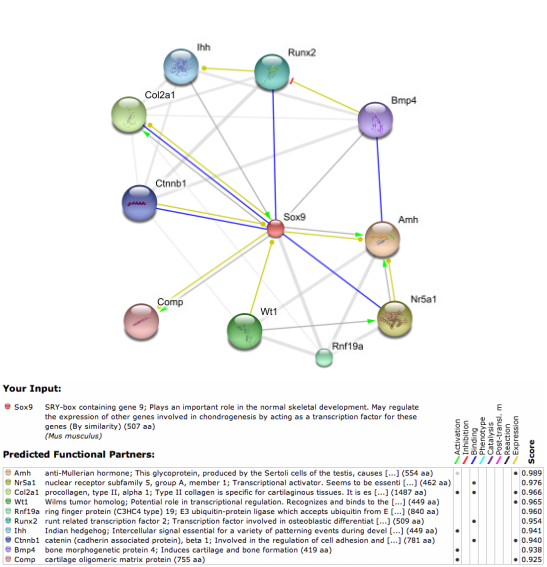
(Mice)
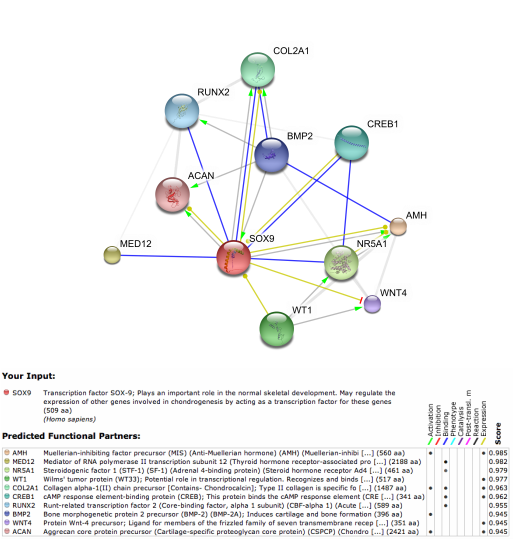
(Humans)
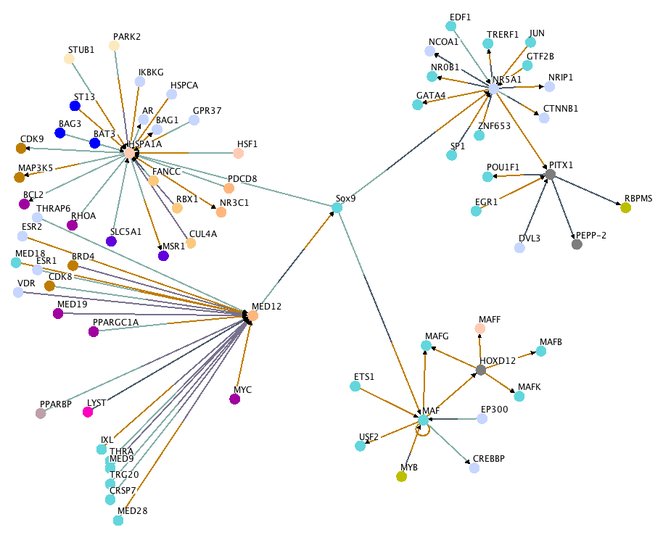
Osprey PPI network with human Sox9
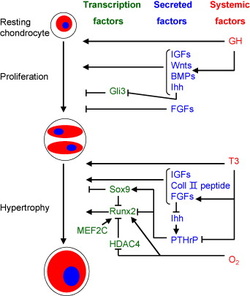
Figure 3: Schematic diagram providing an overview of the roles of systemic factors, locally secreted factors and transcription factors in the regulation of chondrocyte proliferation and hypertrophy during endochondral ossification, which is to put a lot of the interactome data found above into perspective of what has already been found. (Figure taken from Mackie et al 2008)
References
Davis AP, Capecchi MR. (1996) A mutational analysis of the 5' HoxD genes: dissection of genetic interactions during limb development in the mouse. Development. 122(4):1175-85.
Zákány J, Duboule D (1996) Synpolydactyly in mice with a targeted deficiency in the HoxD complex. Nature. 384(6604):69-71.
Kioussi C, Briata P, Baek SH, Rose DW, Hamblet NS, Herman T, Ohgi KA, Lin C, Gleiberman A, Wang J, Brault V, Ruiz-Lozano P, Nguyen HD, Kemler R, Glass CK, Wynshaw-Boris A, Rosenfeld MG. (2002) Identification of a Wnt/Dvl/beta-Catenin --> Pitx2 pathway mediating cell-type-specific proliferation during development. Cell. 111(5):673-85.
Huelsken J, Vogel R, Brinkmann V, Erdmann B, Birchmeier C, Birchmeier W. (2000) Requirement for beta-catenin in anterior-posterior axis formation in mice. J Cell Biol. 148(3):567-78.
Shikama N, Lutz W, Kretzschmar R, Sauter N, Roth JF, Marino S, Wittwer J, Scheidweiler A, Eckner R. (2003) Essential function of p300 acetyltransferase activity in heart, lung and small intestine formation. EMBO J. 22(19):5175-85.
Elliott AM, de Miguel MP, Rebel VI, Donovan PJ. (2007) Identifying genes differentially expressed between PGCs and ES cells reveals a role for CREB-binding protein in germ cell survival. Dev Biol. 311(2):347-58.
Lanctôt C, Moreau A, Chamberland M, Tremblay ML, Drouin J. (1999) Hindlimb patterning and mandible development require the Ptx1 gene. Development. 126(9):1805-10.
De Santa Barbara P, Bonneaud N, Boizet B, Desclozeaux M, Moniot B, Sudbeck P, Scherer G, Poulat F, Berta P. Direct interaction of SRY-related protein SOX9 and steroidogenic factor 1 regulates transcription of the human anti-Müllerian hormone gene. Mol Cell Biol. 1998 Nov;18(11):6653-65.
Tremblay JJ, Marcil A, Gauthier Y, Drouin J. Ptx1 regulates SF-1 activity by an interaction that mimics the role of the ligand-binding domain. EMBO J. 1999 Jun 15;18(12):3431-41.
Day TF, Guo X, Garrett-Beal L, Yang Y. Wnt/beta-catenin signaling in mesenchymal progenitors controls osteoblast and chondrocyte differentiation during vertebrate skeletogenesis. Dev Cell. 2005 May;8(5):739-50.
Hill TP, Später D, Taketo MM, Birchmeier W, Hartmann C. Canonical Wnt/beta-catenin signaling prevents osteoblasts from differentiating into chondrocytes. Dev Cell. 2005 May;8(5):727-38.
Mackie EJ, Ahmed YA, Tatarczuch L, Chen KS, Mirams M. Endochondral ossification: how cartilage is converted into bone in the developing skeleton. Int J Biochem Cell Biol. 2008;40(1):46-62. Epub 2007 Jun 29.
Zákány J, Duboule D (1996) Synpolydactyly in mice with a targeted deficiency in the HoxD complex. Nature. 384(6604):69-71.
Kioussi C, Briata P, Baek SH, Rose DW, Hamblet NS, Herman T, Ohgi KA, Lin C, Gleiberman A, Wang J, Brault V, Ruiz-Lozano P, Nguyen HD, Kemler R, Glass CK, Wynshaw-Boris A, Rosenfeld MG. (2002) Identification of a Wnt/Dvl/beta-Catenin --> Pitx2 pathway mediating cell-type-specific proliferation during development. Cell. 111(5):673-85.
Huelsken J, Vogel R, Brinkmann V, Erdmann B, Birchmeier C, Birchmeier W. (2000) Requirement for beta-catenin in anterior-posterior axis formation in mice. J Cell Biol. 148(3):567-78.
Shikama N, Lutz W, Kretzschmar R, Sauter N, Roth JF, Marino S, Wittwer J, Scheidweiler A, Eckner R. (2003) Essential function of p300 acetyltransferase activity in heart, lung and small intestine formation. EMBO J. 22(19):5175-85.
Elliott AM, de Miguel MP, Rebel VI, Donovan PJ. (2007) Identifying genes differentially expressed between PGCs and ES cells reveals a role for CREB-binding protein in germ cell survival. Dev Biol. 311(2):347-58.
Lanctôt C, Moreau A, Chamberland M, Tremblay ML, Drouin J. (1999) Hindlimb patterning and mandible development require the Ptx1 gene. Development. 126(9):1805-10.
De Santa Barbara P, Bonneaud N, Boizet B, Desclozeaux M, Moniot B, Sudbeck P, Scherer G, Poulat F, Berta P. Direct interaction of SRY-related protein SOX9 and steroidogenic factor 1 regulates transcription of the human anti-Müllerian hormone gene. Mol Cell Biol. 1998 Nov;18(11):6653-65.
Tremblay JJ, Marcil A, Gauthier Y, Drouin J. Ptx1 regulates SF-1 activity by an interaction that mimics the role of the ligand-binding domain. EMBO J. 1999 Jun 15;18(12):3431-41.
Day TF, Guo X, Garrett-Beal L, Yang Y. Wnt/beta-catenin signaling in mesenchymal progenitors controls osteoblast and chondrocyte differentiation during vertebrate skeletogenesis. Dev Cell. 2005 May;8(5):739-50.
Hill TP, Später D, Taketo MM, Birchmeier W, Hartmann C. Canonical Wnt/beta-catenin signaling prevents osteoblasts from differentiating into chondrocytes. Dev Cell. 2005 May;8(5):727-38.
Mackie EJ, Ahmed YA, Tatarczuch L, Chen KS, Mirams M. Endochondral ossification: how cartilage is converted into bone in the developing skeleton. Int J Biochem Cell Biol. 2008;40(1):46-62. Epub 2007 Jun 29.
